Пречишћавање протеина је процес увођења секвенци нуклеинских киселина које кодирају протеине у ћелије домаћина путем техника генетског инжењеринга, узрокујући њихову експресију у великим количинама, а затим их пречишћавају ин витро користећи одговарајуће методе пречишћавања како би се добили протеини високе чистоће, активности и приноса . Системи експресије протеина се могу категорисати у прокариотске системе и еукариотске системе.
Принципи и методе за изазивање експресије протеина у прокариотским системима и пречишћавање протеина ин витро коришћењем афинитетне хроматографије су описани у наставку.
ПРИНЦИП
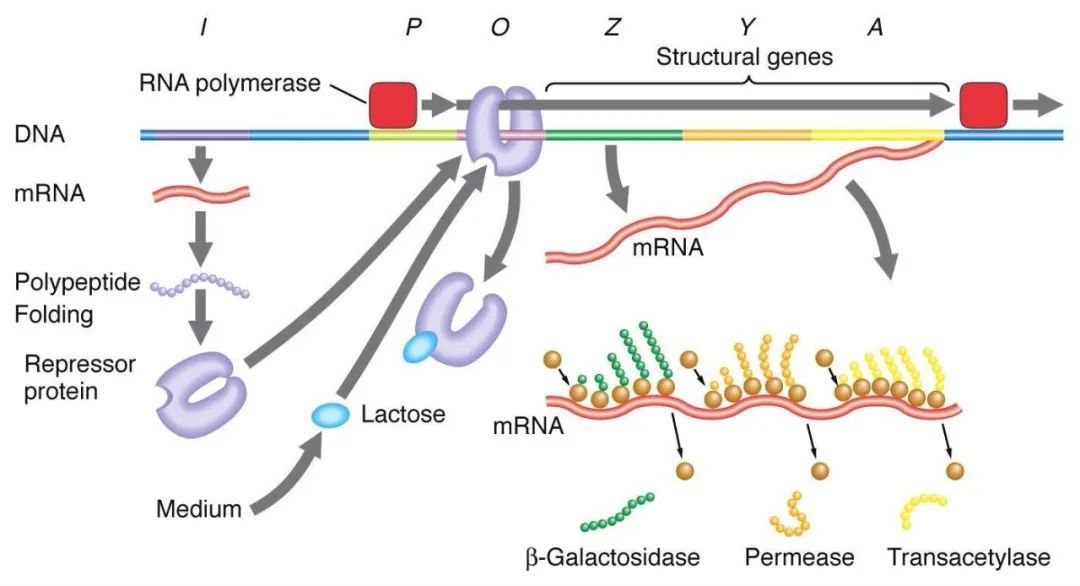
1、Негативна регулација манипулатора лактозе:
У одсуству лактозе, лац манипулатор је у стању одвраћања, у то време И секвенца експримира протеин одвраћања Лац да се веже за О секвенцу, спречавајући РНК полимеразу да се веже за П секвенцу и инхибира иницијацију транскрипције. Када је лактоза присутна, лактоза улази у ћелију и катализује је β-галактозидаза и претвара се у изолактозу, која се везује за инхибиторни протеин, изазивајући конформациону промену у протеину и доводећи до дисоцијације инхибиторног протеина из О секвенце, чиме се покреће транскрипција.
Изопропилтиогалактозид (ИПТГ) делује на исти начин као изогалактоза и изузетно је моћан индуктор који се не метаболише од стране бактерија и веома је стабилан, те се стога широко користи у лабораторијама.
2、Одабир соја домаћина:
(1) БЛ21 је један од најчешће коришћених сојева за прокариотску експресију, који је углавном погодан за експресију нетоксичних протеина са Е. цоли полимеразом, па се може применити на експресију прокариотских система коришћењем Е. цоли РНК полимеразе као што су тац или трц (нпр. пГЕКС, пМАЛ плазмиди).
(2) БЛ21(ДЕ3) интегрише ген Т7 фаг РНК полимеразе у λ фаг ДЕ3 регион на хромозому соја БЛ21, који може да експримира и Т7 РНК полимеразу и Е. цоли РНК полимеразу , и може се користити за експресију плазмида и тако на пЕТ серије, пЕТ серије, пГ
3, методе пречишћавања протеина:
(1) гел филтрациона хроматографија: према величини молекула из мешавине у одвојене протеине, облику различитих протеина и молекуларној величини разлика у смеши кроз колону гел филтрационе хроматографије која садржи честице пунила, због тога што је молекуларна величина различитих протеина различита, дифузија у специфичну величину отвора отвора за честице првог молекула ће бити велике честице протеина. који се елуирају из молекула су мањи што је касније елуција.
(2) Хроматографија јонске измене: раздвајање и пречишћавање протеина засновано је на различитим набојима на површини протеина, површина протеина је обично једнолико наелектрисана и може се комбиновати са колонама за катјоне/ањонске измене под одређеним условима. Када се пХ промени или се за елуирање користи пуфер са постепено растућом јонском снагом, везана супстанца може да се размени са јонима у елуенту и елуира у раствор. Пошто различите супстанце имају различита наелектрисања и различите способности везивања са колоном за измену јона, редослед елуирања у раствор је такође различит.
(3) Хидрофобна хроматографија: Користећи хидрофобност протеина, хидрофобни остаци ће бити изложени на површини протеина након денатурације или у окружењу са високим садржајем соли, хидрофобни остаци различитих протеина имају различиту јачину дејства са хидрофобним лигандима стационарне фазе, а хидрофобност најслабије компоненте може се користити као најјаче компоненте иопарације. елуент по реду од високог ка ниском.
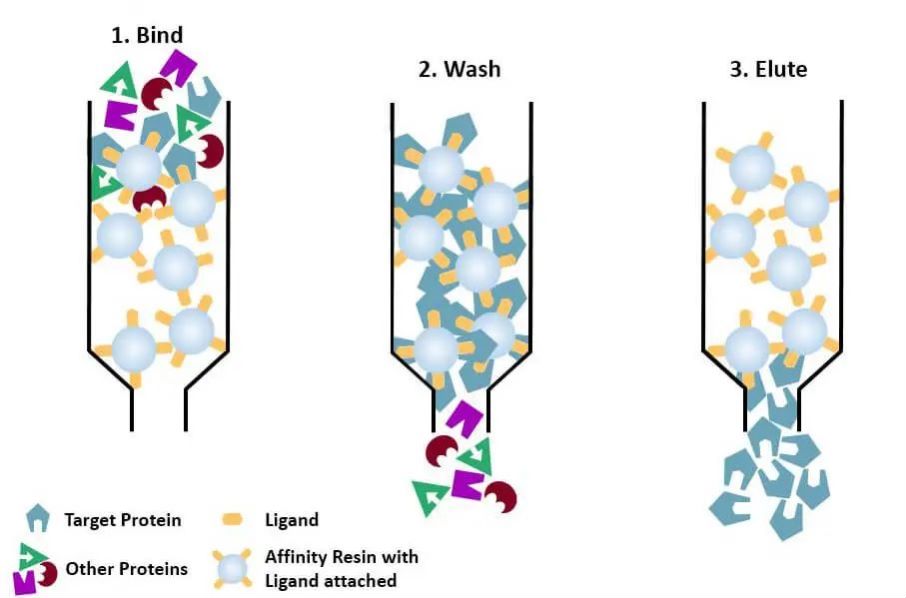
(4) Афинитетна хроматографија: специфични лиганд протеина који треба да се пречисти (или обележен на протеину) је ковалентно везан за молекул носача одговарајућим хемијским методама. Када се мешавина протеина дода у хроматографску колону испуњену афинитетним медијумом, протеин који се пречишћава је специфично везан за лиганд, док се остали протеини не везују и уклањају испирањем, а специфично везани протеин може да се елуира раствором слободног одговарајућег лиганда. Специфично везани протеини могу бити елуирани раствором који садржи одговарајући одговарајући лиганд.
4、Избор ознака за пречишћавање протеина:
Карактеристике различитих ознака су следеће↓
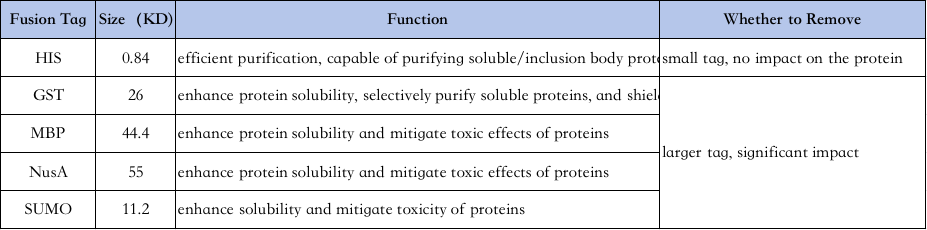
Пречишћавање Хис-таг-а: Хис-таг је један од најчешће коришћених ознака, где се 6-10 хистидина додаје амино или карбоксилном крају протеина, а циљни протеин се пречишћава својом способношћу да се чврсто везује за Ни2+ хелатне колоне у нормалним или денатурационим условима (нпр. 8М уреа), а затим се елуира у амино или карбоксил-амино аминокиселине. бунари за перле).
Инструменти и потрошни материјал
Лабораторијски реагенси:
ИПТГ; ЛБ медиум;
Пуфер за лизу: 20 мМ Трис (7.9), 500 мМ НаЦл, 10 мМ Имидазол;
Пуфер за испирање: 20 мМ Трис (7.9), 500 мМ НаЦл, 20 мМ Имидазол;
Пуфер за елуирање: 20 мМ Трис (7.9), 200 мМ НаЦл, 300 мМ имидазола ( концентрација имидазола се може подесити према ефикасности елуирања );
Раствор за дијализу: 20 мМ Трис (7.9), 50 мМ НаЦл, 10% глицерол;
Каомас Бриллиант Блуе раствор за бојење; раствор за деколоризацију
Инструменти:
Ни-НТА, ултразвучни разбијач, хроматографска колона, концентрационе цеви

Процедура
1、Конструкција прокариотског експресионог плазмида: ПЦР циљног гена, векторска дигестија, лигација, трансформација, одабир појединачног клона за секвенцирање;
2、Трансформишите успешно конструисани плазмид у БЛ21(ДЕ3), инкубирајте на 37℃ преко ноћи и изаберите један клон за мало мућкање преко ноћи;
3、Мала индукција да би се открили најбољи услови индукције: протресаће се преко ноћи бактеријски раствор 1:1000 инокулација у 3мЛ ЛБ, 37 ℃ мућкати 4-5х до бактеријског раствора ОД600 = 0,6-0,8, додати различите концентрације ИПТГ-а, време ниже температуре (0,1-1), ниже температуре продужено, као што је 37 ℃ индуковано 4-5 х, 30 ℃ индуковано 6 х-8 х, 16 ℃ индуковано 16-20 х, 16 ℃ индуковано 16-20 х, температура се смањује, време се продужава, као што је 37 ℃-5, индуковано 37 ℄ 5 х, индуковано 3 х-6 х 16°Ц индуковано 16-20х, температура се смањује. Ц током 16-20 х, узмите бактеријски сок пре и после индукције у различитим условима индукције, пустите гел, обојите и посматрајте резултате индукције; (Уопштено говорећи, што је нижа концентрација ИПТГ и што је нижа температура индукције, спорија је експресија циљног протеина, то је погодније за правилно савијање протеина, чиме се повећава њихова растворљивост и смањује стварање инклузијских тела)
4、Након проналажења најпогоднијих услова за индукцију, инокулирајте раствор бактерије 1:1000 у 2Л ЛБ, протресите на 37℃ док ОД600 бактеријског раствора=0,6-0,8, аспирирајте 20μЛ раствора бактерије и оставите да ради гел добијени протеин у одговарајућим условима (1), а затим у условима за индукцију добијеног протеина пре-експериментишите, аспирирајте 20 μЛ раствора бактерије и оставите да тече гел (2);
5. Уклоните индуковану бактеријску течност, центрифугирајте на 4000 г 15 минута на 4℃;
6、Одбаците супернатант, измерите, додајте 10 мЛ пуфера за лизу (1:100 плус инхибитор протеазе) за сваки грам бактерије, ресуспендујте, ставите на лед око 30 минута;
7、Грушење под притиском: одложите алкохол у хомогенизатору високог притиска, исперите водом два пута, користите пуфер за лизу да бисте уравнотежили једном, додајте бактеријску течност, притисак (притисак не би требало да прелази 800 кпа), бактеријска течност преко три до пет пута до провидне и нелепљиве;
8. Сакупите згњечену бактеријску течност, 12000 г, центрифугирање на 4 ℃ током 20 минута, одвајање супернатанта и талога, сваки је задржао узорак од 20 μЛ (3) (4) да би се покренуо гел;
9、2мЛ Ни-НТА је додато у колону за пречишћавање, да би се филтрирао етанол, испрао водом и додат је пуфер за лизу да би се колона уравнотежила;
10、Са ресуспензијом Ни-НТА пуфера за лизу додат у супернатант, добро промешати, 4 ℃ инкубирати у шејкеру током 2 х;
11、Супернатант је пропуштен кроз колону на 4°Ц и сакупљено је 20 μЛ филтрата (5);
12、Исперите колону са 5мЛ пуфера за испирање 3 пута, сакупите узорак филтрата 20μЛ (6);
13、Додајте 1мЛ пуфера за елуирање, инкубирајте 5мин, сакупите елуат, поновите 5 пута, сакупите 5мЛ елуата у исту епрувету, оставите 20μЛ узорка (7);
14、Појединачни узорци протеина добијени током експеримента су пуштени, обојени Цаумас Бриллиант Блуе током 1 х, и обезбојени све док плава боја позадине није светлија и јасне протеинске траке нису биле видљиве;
15、Анализирајте протеинске траке сваког узорка, према резултатима накнадне операције: ако елуирани узорци протеина у протеину без хетерозиготних трака или хетерозиготних трака или мање и плитки, могу бити дијализа и концентрација, ако више хетерозиготних трака треба поново да се пречисти након дијализе и концентрације.
16、Дијализа: додајте узорак у мембрану за дијализу, стегните оба краја, дијализа на 4℃ преко ноћи;
17、Концентрација: према величини молекулске тежине узорка да се изабере одговарајућа епрувета за концентрацију 4 ℃ концентрације ниске брзине, концентрација мерења концентрације протеина, обележавање, у брзом замрзавању течног азота, а затим замрзнуто у фрижидеру -80 ℃.
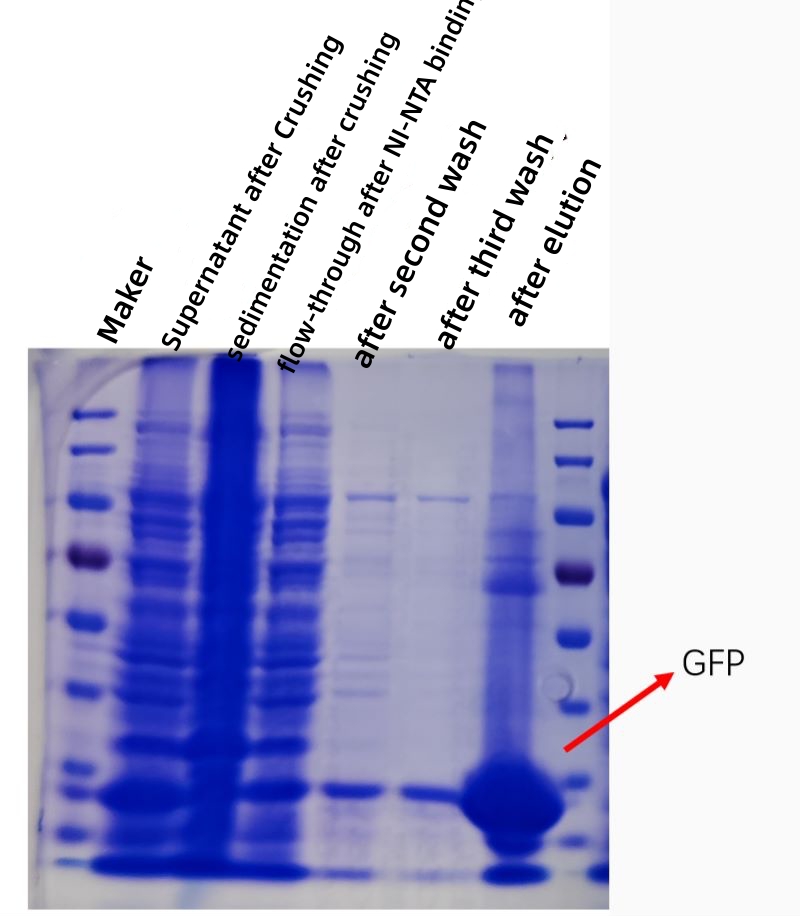
РЕЗУЛТАТ
Слика испод приказује кофекциони дијаграм пречишћавања афинитета ГФП протеина ин витро, а чистоћа и концентрација пречишћеног протеина су знатно повећане