Eiwitzuivering is het proces waarbij nucleïnezuursequenties die coderen voor eiwitten in gastheercellen worden geïntroduceerd door middel van genetische manipulatietechnieken, waardoor ze in grote hoeveelheden tot expressie worden gebracht, en ze vervolgens in vitro worden gezuiverd met behulp van geschikte zuiveringsmethoden om eiwitten met een hoge zuiverheid, activiteit en opbrengst te verkrijgen . Eiwitexpressiesystemen kunnen worden onderverdeeld in prokaryotische systemen en eukaryotische systemen.
De principes en methoden voor het induceren van eiwitexpressie in prokaryotische systemen en het in vitro zuiveren van eiwitten met behulp van affiniteitschromatografie worden hieronder beschreven.
BEGINSEL
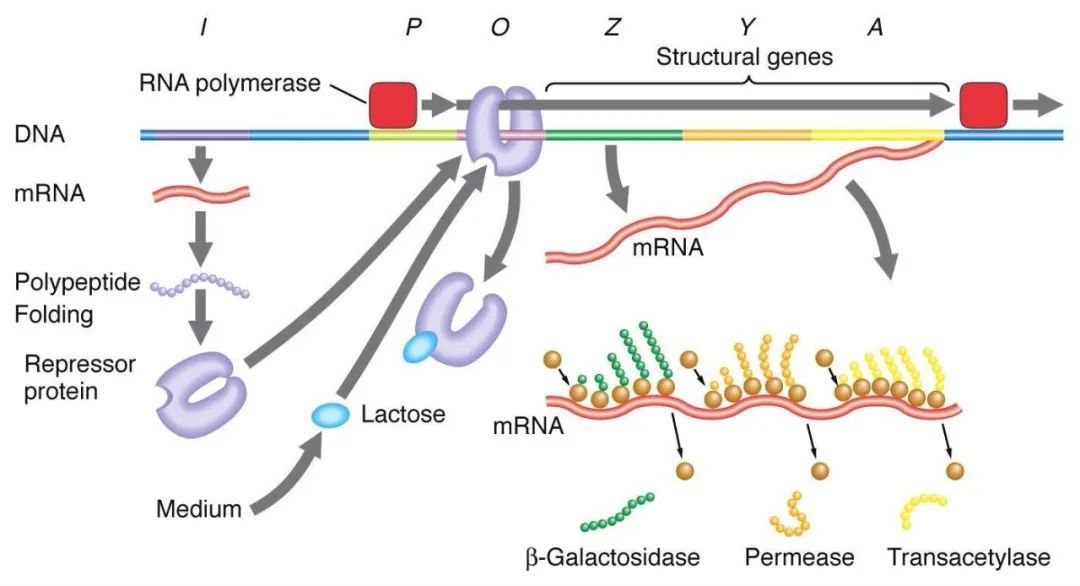
1, negatieve regulatie van de lactosemanipulator:
Bij afwezigheid van lactose bevindt de lac-manipulator zich in een staat van afschrikking, op welk moment de I-sequentie het Lac-afschrikkende eiwit tot expressie brengt om aan de O-sequentie te binden, waardoor wordt voorkomen dat RNA-polymerase aan de P-sequentie bindt en de transcriptie-initiatie wordt geremd. Wanneer lactose aanwezig is, komt lactose de cel binnen en wordt gekatalyseerd door β-galactosidase en omgezet in isolactose, dat zich bindt aan het remmende eiwit, waardoor een conformationele verandering in het eiwit ontstaat en leidt tot dissociatie van het remmende eiwit van de O-sequentie, waardoor transcriptie wordt geïnitieerd.
Isopropylthiogalactoside (IPTG) werkt op dezelfde manier als isogalactose en is een uiterst krachtige inductor die niet door bacteriën wordt gemetaboliseerd en zeer stabiel is, en wordt daarom veel gebruikt in laboratoria.
2. Selectie van gastheerstammen:
(1) BL21 is een van de meest gebruikte stammen voor prokaryotische expressie, die voornamelijk geschikt is voor de expressie van niet-toxische eiwitten met E. coli-polymerase, zodat het kan worden toegepast op de expressie van prokaryotische systemen met behulp van E. coli-RNA-polymerase zoals tac of trc (bijv. pGEX, pMAL-plasmiden).
(2) BL21(DE3) integreert het T7-faag-RNA-polymerasegen in het λ-faag DE3-gebied op het chromosoom van de BL21-stam, dat zowel T7-RNA-polymerase als E. coli-RNA-polymerase tot expressie kan brengen , en kan worden gebruikt voor de expressie van plasmiden zoals de pET-serie, pGEX, pMAL, enzovoort.
3, eiwitzuiveringsmethoden:
(1) Gelfiltratiechromatografie: afhankelijk van de moleculaire grootte van het mengsel om eiwitten te scheiden, de vorm van verschillende eiwitten en de moleculaire grootte van de verschillen in het mengsel door de gelfiltratiechromatografiekolom die vulstofdeeltjes bevat, vanwege de moleculaire grootte van verschillende eiwitten zijn verschillend, de diffusie in de specifieke grootte van de openingdeeltjes van het vermogen om te variëren, hoe groter de eiwitmoleculen de eerste zullen zijn die uit de moleculen worden geëlueerd, hoe later hoe later de elutie.
(2) Ionenuitwisselingschromatografie: eiwitscheiding en -zuivering is gebaseerd op de verschillende ladingen op het oppervlak van het eiwit, het oppervlak van het eiwit is gewoonlijk uniform geladen en kan onder bepaalde omstandigheden worden gecombineerd met kation/anion-uitwisselingskolommen. Wanneer de pH wordt veranderd of een buffer met geleidelijk toenemende ionensterkte wordt gebruikt voor elutie, kan de gebonden stof worden uitgewisseld met de ionen in het eluens en in de oplossing worden geëlueerd. Omdat verschillende stoffen verschillende ladingen hebben en verschillende bindingsmogelijkheden hebben met de ionenuitwisselingskolom, is de volgorde waarin ze in oplossing worden geëlueerd ook anders.
(3) Hydrofobe chromatografie: met behulp van de hydrofobiciteit van eiwitten zullen hydrofobe residuen worden blootgesteld aan het oppervlak van eiwitten na denaturatie of in een omgeving met een hoog zoutgehalte. De hydrofobe residuen van verschillende eiwitten hebben verschillende werkingssterkten met de hydrofobe liganden van de stationaire fase, en hydrofobiciteit kan worden gebruikt als de zwakste tot de sterkste componentscheiding door de ionsterkte van het eluens te gebruiken in de volgorde van hoog naar laag.
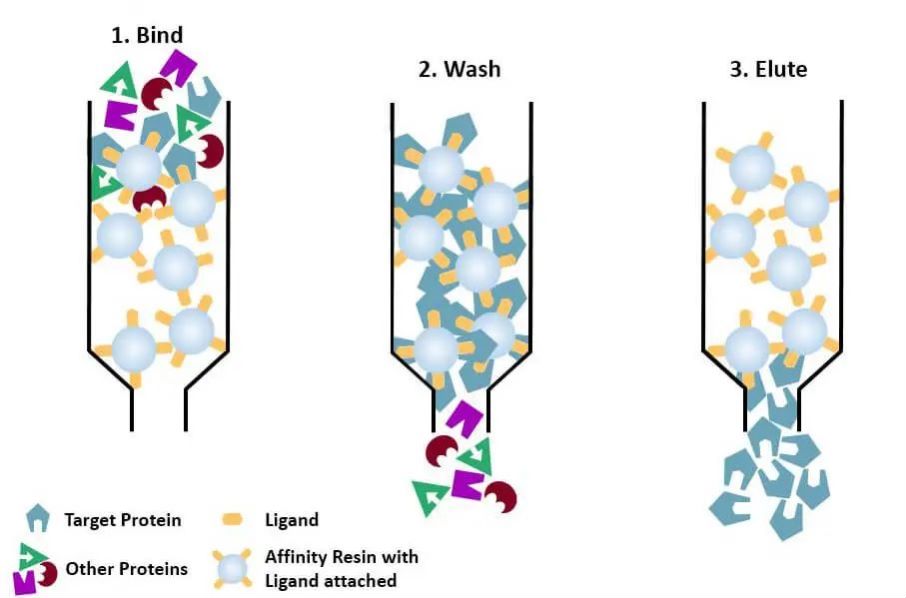
(4) Affiniteitschromatografie: het specifieke ligand van een te zuiveren eiwit (of te taggen op het eiwit) wordt door geschikte chemische methoden covalent aan het dragermolecuul gebonden. Wanneer het eiwitmengsel wordt toegevoegd aan de chromatografische kolom gevuld met affiniteitsmedium, wordt het te zuiveren eiwit specifiek gebonden aan het ligand, terwijl de andere eiwitten niet worden gebonden en verwijderd door wassen, en kan het specifiek gebonden eiwit worden geëlueerd met een oplossing van het vrije overeenkomstige ligand. De specifiek gebonden eiwitten kunnen worden geëlueerd met een oplossing die het vrije overeenkomstige ligand bevat.
4. Selectie van eiwitzuiveringslabels:
De kenmerken van verschillende labels zijn als volgt↓
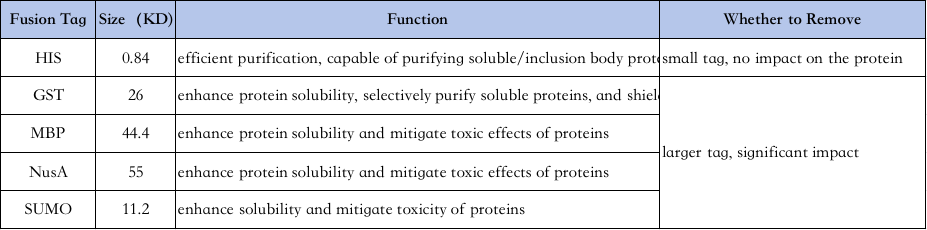
His-tag-zuivering: His-tag is een van de meest gebruikte tags, waarbij 6-10 histidine wordt toegevoegd aan het amino- of carboxyl-uiteinde van het eiwit, en het doeleiwit wordt gezuiverd door zijn vermogen om stevig te binden aan Ni2+-chelerende kolommen onder normale of denaturerende omstandigheden (bijv. 8M ureum), en vervolgens geëlueerd met imidazol (dat concurreert om binding aan nikkelionen in de parelputten).
Instrumenten en verbruiksartikelen
Laboratoriumreagentia:
IPTG; LB-medium;
Lysisbuffer: 20 mM Tris (7,9), 500 mM NaCl, 10 mM imidazool;
Wasbuffer: 20 mM Tris (7,9), 500 mM NaCl, 20 mM imidazool;
Elutiebuffer: 20 mM Tris (7.9), 200 mM NaCl, 300 mM imidazool ( de imidazoolconcentratie kan worden aangepast aan de elutie-efficiëntie );
Dialyseoplossing: 20 mM Tris (7,9), 50 mM NaCl, 10% glycerol;
Kaomas Brilliant Blue-kleuringsoplossing; ontkleurende oplossing
Instrumenten:
Ni-NTA, ultrasone breker, chromatografiekolom, concentratiebuizen

Procedure
1. Constructie van prokaryotisch expressieplasmide: PCR van doelgen, vectordigestie, ligatie, transformatie, het kiezen van een enkele kloon voor sequencing;
2. Transformeer het met succes geconstrueerde plasmide in BL21 (DE3), incubeer een nacht bij 37 ℃ en kies de enkele kloon voor een nachtje schudden;
3. Kleine inductie om de beste inductieomstandigheden te vinden: zal 's nachts bacteriële oplossing 1:1000 inenting worden geschud in 3 ml LB, 37 ℃ 4-5 uur schudden met de bacteriële oplossing OD600 = 0,6-0,8, verschillende IPTG-concentraties toevoegen (0,1-1 mM), verschillende temperaturen, met het verlagen van de temperatuur wordt de inductietijd verlengd, zoals 37 ℃ geïnduceerd 4-5 uur, 30 ℃ geïnduceerd 6 uur-8 uur, 16 ℃ geïnduceerd 16-20 uur, 16 ° C geïnduceerd 16-20 uur, de temperatuur wordt verlaagd, de tijd wordt verlengd, zoals 37 ℃ geïnduceerd 4-5 uur, 30 ℃ geïnduceerd 6 uur-8 uur, 16 ° C geïnduceerd 16-20 uur, de temperatuur is verminderd. C gedurende 16-20 uur, neem het bacteriesap voor en na inductie onder verschillende inductieomstandigheden, laat de gel lopen, kleur en observeer de resultaten van inductie; (Over het algemeen geldt: hoe lager de IPTG-concentratie en hoe lager de inductietemperatuur, hoe langzamer de expressie van het doeleiwit, hoe bevorderlijker voor de juiste vouwing van eiwitten, waardoor hun oplosbaarheid toeneemt en de vorming van inclusielichamen wordt verminderd)
4. Na het vinden van de meest geschikte inductieomstandigheden, ent u de bacteriële oplossing 1:1000 in 2 liter LB, schudt u bij 37 ℃ tot de OD600 van de bacteriële oplossing = 0,6-0,8, zuigt u 20 μl van de bacteriële oplossing op en laat u deze de gel laten lopen (1), induceer vervolgens de eiwitexpressie onder de geschikte inductieomstandigheden verkregen in het pre-experiment, en zuig 20 μl op van de bacteriële oplossing en laat de gel lopen (2);
5. Verwijder de geïnduceerde bacteriële vloeistof, centrifugeer bij 4000 g gedurende 15 minuten bij 4 ℃;
6. Gooi het supernatant weg, weeg, voeg 10 ml lysisbuffer (1:100 plus proteaseremmer) toe voor elke gram bacteriën, suspendeer opnieuw en plaats ongeveer 30 minuten op ijs;
7. Druk verpletteren: zet de alcohol in de hogedrukhomogenisator uit, spoel twee keer met water, gebruik Lysisbuffer om één keer in evenwicht te brengen, voeg de bacteriële vloeistof toe, druk opbrengen (de druk mag niet hoger zijn dan 800 kpa), de bacteriële vloeistof meer dan drie tot vijf keer tot transparant en niet plakkerig;
8. Verzamel de gemalen bacteriële vloeistof, 12.000 g, 4 ℃ centrifugatie gedurende 20 minuten, de supernatant en neerslagscheiding, elk behield 20 μL monster (3) (4) om gel te laten lopen;
9,2 ml Ni-NTA werd aan de zuiveringskolom toegevoegd, om met ethanol te worden gefilterd, gespoeld met water en er werd lysisbuffer toegevoegd om de kolom in evenwicht te brengen;
10. Met Lysisbuffer Ni-NTA-resuspensie toegevoegd aan het supernatant, goed mengen, incubatie in een schudapparaat van 4 ℃ gedurende 2 uur;
11. Het supernatant werd bij 4°C door de kolom geleid en 20 μl filtraat werd verzameld (5);
12. Was de kolom 3 keer met 5 ml wasbuffer, verzamel het filtraatmonster 20 μL (6);
13. Voeg 1 ml eluaatbuffer toe, incubeer gedurende 5 minuten, verzamel het eluaat, herhaal 5 keer, verzamel 5 ml eluaat in dezelfde buis, laat 20 μl monster achter (7);
14. De individuele eiwitmonsters die in de loop van het experiment werden verkregen, werden onderzocht, gedurende 1 uur gekleurd met Caumas Brilliant Blue en ontkleurd totdat de blauwe achtergrondkleur lichter was en heldere eiwitbanden zichtbaar waren;
15. Analyseer de eiwitbanden van elk monster, volgens de resultaten van de daaropvolgende operatie: als de geëlueerde eiwitmonsters in het eiwit zonder heterozygote banden of heterozygote banden of minder en ondiep zijn, dialyse en concentratie kunnen zijn, als meer heterozygote banden opnieuw moeten worden gezuiverd na dialyse en concentratie.
16. Dialyse: voeg het monster toe aan het dialysemembraan, klem beide uiteinden vast, dialyse bij 4 ℃ gedurende de nacht;
17, Concentratie: afhankelijk van de molecuulgewichtsgrootte van het monster, kies de juiste concentratiebuis 4 ℃ lage snelheidsconcentratie, concentratie van eiwitconcentratiemeting, etikettering, in vloeibare stikstof snel invriezen en vervolgens ingevroren in -80 ℃ koelkast.
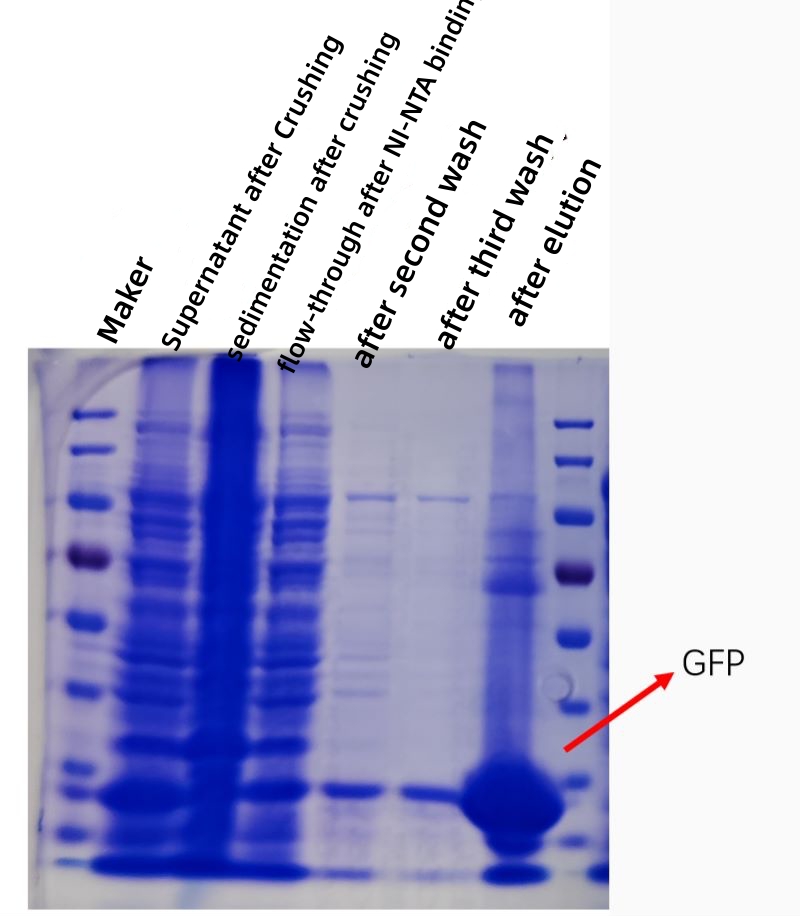
RESULTAAT
De onderstaande figuur toont een kofectiegrafiek van GFP-eiwitaffiniteitszuivering in vitro, en de zuiverheid en concentratie van het gezuiverde eiwit is aanzienlijk toegenomen