Proteinrensing er prosessen med å introdusere nukleinsyresekvenser som koder for proteiner i vertsceller gjennom genteknologiske teknikker, få dem til å uttrykkes i store mengder, og deretter rense dem in vitro ved å bruke passende rensemetoder for å oppnå proteiner med høy renhet, aktivitet og utbytte . Proteinekspresjonssystemer kan kategoriseres i prokaryote systemer og eukaryote systemer.
Prinsippene og metodene for å indusere proteinekspresjon i prokaryote systemer og rense proteiner in vitro ved bruk av affinitetskromatografi er beskrevet nedenfor.
PRINSIPP
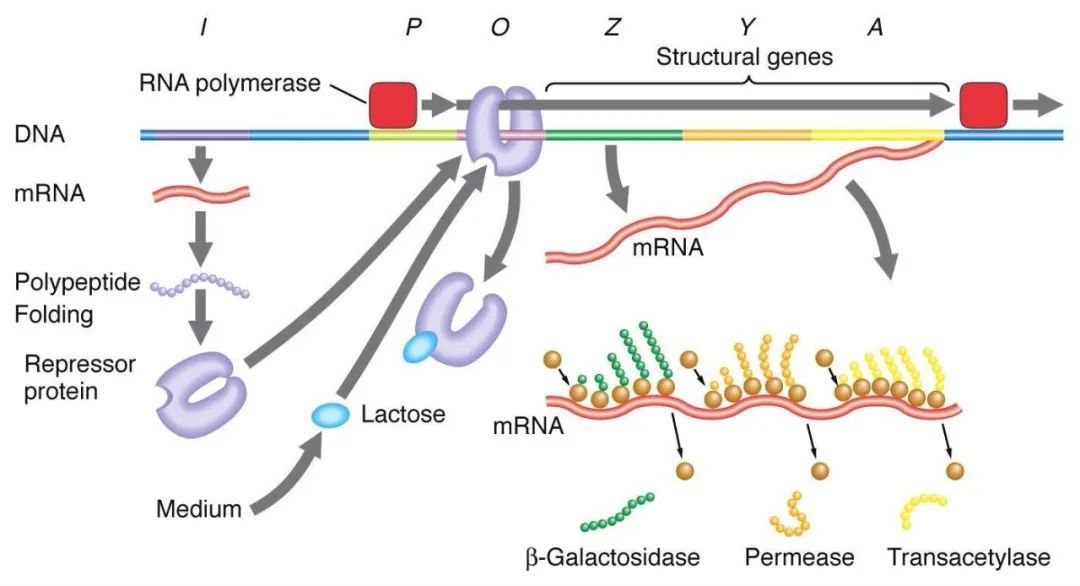
1, Negativ regulering av laktosemanipulatoren:
I fravær av laktose er lac-manipulatoren i en avskrekkingstilstand, på hvilket tidspunkt I-sekvensen uttrykker Lac-avskrekkende protein for å binde seg til O-sekvensen, forhindre RNA-polymerase i å binde seg til P-sekvensen og hemme transkripsjonsinitiering. Når laktose er tilstede, kommer laktose inn i cellen og katalyseres av β-galaktosidase og omdannes til isolaktose, som binder seg til det inhiberende proteinet, noe som forårsaker en konformasjonsendring i proteinet og fører til dissosiasjon av det hemmende proteinet fra O-sekvensen, og initierer derved transkripsjon.
Isopropyltiogalaktosid (IPTG) virker på samme måte som isogalaktose og er en ekstremt potent induktor som ikke metaboliseres av bakterier og er meget stabil, og er derfor mye brukt i laboratorier.
2、valg av vertsstamme:
(1) BL21 er en av de mest brukte stammene for prokaryot ekspresjon, som hovedsakelig er egnet for ekspresjon av ikke-toksiske proteiner med E. coli polymerase, så den kan brukes på ekspresjon av prokaryote systemer ved bruk av E. coli RNA polymerase som tac eller trc (f.eks. pGEX, pMAL plasmider).
(2) BL21(DE3) integrerer T7-fag-RNA-polymerase-genet i λ-fag DE3-regionen på kromosomet til BL21-stammen, som kan uttrykke både T7-RNA-polymerase og E. coli RNA-polymerase , og kan brukes for ekspresjon av plasmider som pET-serien, pGEX, og so on.MAL
3, proteinrensingsmetoder:
(1) Gelfiltreringskromatografi: i henhold til molekylstørrelsen fra blandingen til å separere proteiner, formen til forskjellige proteiner og molekylstørrelsen til forskjellene i blandingen gjennom gelfiltreringskromatografikolonnen som inneholder fyllpartikler, på grunn av molekylstørrelsen til forskjellige proteiner er forskjellige, vil diffusjonen til den spesifikke størrelsen til åpningspartikkelen være større, molekylene til åpningspartikkelen varierer. først som skal elueres ut av molekylene er mindre jo senere jo senere elueringen.
(2) Ionebytterkromatografi: proteinseparasjon og rensing er basert på de forskjellige ladningene på overflaten av proteinet, overflaten av proteinet er vanligvis jevnt ladet, og kan kombineres med kation/anionbytterkolonner under visse forhold. Når pH endres eller en buffer med gradvis økende ionestyrke brukes til eluering, kan det bundne stoffet byttes ut med ionene i elueringsmidlet og elueres inn i løsningen. Siden forskjellige stoffer har forskjellige ladninger og forskjellige bindingsevner med ionebytterkolonnen, er rekkefølgen for eluering i løsning også forskjellig.
(3) Hydrofob kromatografi: Ved bruk av hydrofobiciteten til proteiner vil hydrofobe rester eksponeres på overflaten av proteiner etter denaturering eller i et miljø med høyt saltinnhold, de hydrofobe restene av forskjellige proteiner har forskjellig virkningsstyrke med de hydrofobe ligander i den stasjonære fasen, og hydrofobicitet kan brukes som den svakeste separasjonsstyrken til den sterkeste komponenten. i rekkefølge fra høy til lav.
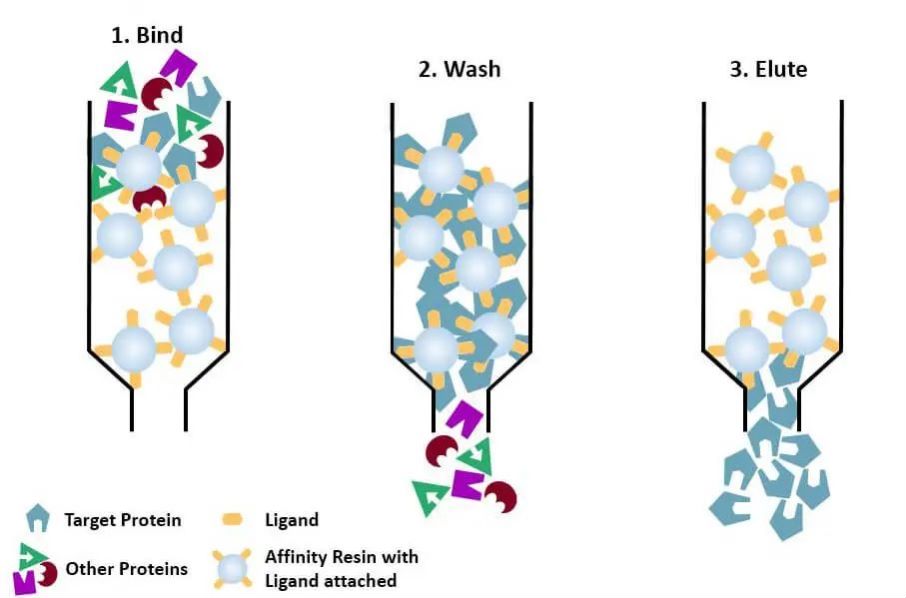
(4) Affinitetskromatografi: den spesifikke liganden til et protein som skal renses (eller merkes på proteinet) festes kovalent til bærermolekylet ved hjelp av passende kjemiske metoder. Når proteinblandingen tilsettes til den kromatografiske kolonnen fylt med affinitetsmedium, bindes proteinet som skal renses spesifikt til liganden, mens de andre proteinene ikke bindes og fjernes ved vasking, og det spesifikt bundne proteinet kan elueres av med en løsning av fri tilsvarende ligand. De spesifikt bundne proteinene kan elueres med en løsning som inneholder den frie korresponderende ligand.
4、Utvalg av proteinrenseetiketter:
Egenskapene til forskjellige etiketter er som følger↓
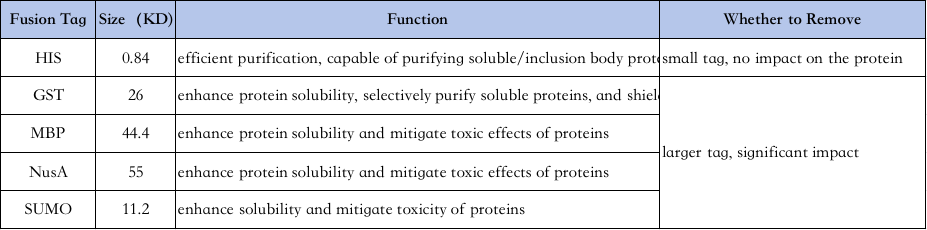
His-tag-rensing: His-tag er en av de mest brukte taggene, der 6-10 histidin tilsettes til amino- eller karboksylterminalen av proteinet, og målproteinet renses ved dets evne til å binde seg tett til Ni2+ chelaterende kolonner under normale eller denaturerende forhold (f.eks. 8M urea), og deretter elueres med imidetazolioner (imidazolioner). perlebrønner).
Instrumenter og forbruksvarer
Laboratoriereagenser:
IPTG; LB medium;
Lyseringsbuffer: 20 mM Tris (7,9), 500 mM NaCl, 10 mM imidazol;
Vaskebuffer: 20 mM Tris (7,9), 500 mM NaCl, 20 mM imidazol;
Elueringsbuffer: 20 mM Tris (7,9), 200 mM NaCl, 300 mM imidazol ( Imidazol-konsentrasjonen kan justeres i henhold til elueringseffektiviteten );
Dialyseløsning: 20 mM Tris (7,9), 50 mM NaCl, 10 % glyserol;
Kaomas Brilliant Blue fargeløsning; avfargingsløsning
Instrumenter:
Ni-NTA, ultralydbryter, kromatografikolonne, konsentrasjonsrør

Prosedyre
1、Konstruksjon av prokaryot ekspresjonsplasmid: PCR av målgenet, vektorfordøyelse, ligering, transformasjon, plukke enkelt klon for sekvensering;
2、Transformer det vellykket konstruerte plasmidet til BL21(DE3), inkuber ved 37 ℃ over natten, og velg enkeltklonen for liten risting over natten;
3、Liten induksjon for å finne ut de beste induksjonsforholdene: vil bli ristet over natten bakteriell løsning 1:1000 inokulering i 3mL LB, 37 ℃ risting 4-5t til bakterieløsningen OD600 = 0,6-0,8, legg til forskjellige konsentrasjoner av IPTG (0,1-1mM forlenget temperatur) slik som 37 ℃ indusert 4-5t, 30 ℃ indusert 6t-8t, 16 ℃ indusert 16-20t, 16 °C indusert 16-20t, temperaturen reduseres, tiden er forlenget, for eksempel 37 ℃ indusert 30-5t, -6 t. ° C indusert 16-20t, temperaturen reduseres. C i 16-20 timer, ta bakteriesaften før og etter induksjon under forskjellige induksjonsforhold, kjør gelen, farg og observer resultatene av induksjonen; (Generelt sett, jo lavere konsentrasjonen av IPTG og jo lavere induksjonstemperaturen er, jo langsommere uttrykket av målproteinet, desto mer bidrar til riktig folding av proteiner, og øker dermed deres løselighet og reduserer genereringen av inklusjonslegemer)
4、Etter å ha funnet de mest egnede induksjonsforholdene, inokuler bakterieløsningen 1:1000 i 2L LB, rist ved 37 ℃ til OD600 av bakterieoppløsningen = 0,6-0,8, aspirer 20μL av bakterieløsningen og la den kjøre i gelen induksjons-proteinet (1), og deretter under egnet induksjonsprotein (1) pre-eksperiment, og aspirer 20μL av bakterieoppløsningen og la den kjøre gelen (2);
5, Fjern den induserte bakterievæsken, sentrifuger ved 4000g i 15 minutter ved 4℃;
6、 Kast supernatanten, vei, tilsett 10mL lyseringsbuffer (1:100 pluss proteasehemmer) for hvert gram bakterier, resuspender, legg på is i ca. 30 minutter;
7、 Trykkknusing: legg av alkoholen i høytrykkshomogenisatoren, skyll med vann to ganger, bruk lyseringsbuffer for å balansere én gang, tilsett bakterievæsken, trykksetting (trykket bør ikke overstige 800kpa), bakterievæsken over tre til fem ganger til gjennomsiktig og ikke klebrig;
8、Samle den knuste bakterievæsken, 12000g, 4 ℃ sentrifugering i 20 minutter, supernatanten og bunnfallseparasjonen, hver beholdt 20μL prøve (3) (4) som skal kjøres gel;
9,2 ml Ni-NTA ble tilsatt rensekolonnen, for å bli filtrert med etanol, skyllet med vann og tilsatt lyseringsbuffer for å ekvilibrere kolonnen;
10、Med lyseringsbuffer Ni-NTA resuspensjon tilsatt til supernatanten, bland godt, 4 ℃ shaker inkubasjon i 2 timer;
11、Supernatanten ble ført gjennom kolonnen ved 4°C, og 20 μL filtrat ble samlet (5);
12、Vask kolonnen med 5mL vaskebuffer 3 ganger, samle filtratprøven 20μL (6);
13、Tilsett 1 mL elueringsbuffer, inkuber i 5 minutter, samle eluatet, gjenta 5 ganger, samle 5 mL eluat i samme rør, la 20 μL prøve (7);
14、De individuelle proteinprøvene som ble oppnådd i løpet av eksperimentet ble kjørt, farget med Caumas Brilliant Blue i 1 time og avfarget inntil den blå bakgrunnsfargen var lysere og klare proteinbånd var synlige;
15、Analyser proteinbåndene til hver prøve, i henhold til resultatene av den påfølgende operasjonen: hvis de eluerte proteinprøvene i proteinet uten heterozygote bånd eller heterozygote bånd eller mindre og grunne, kan være dialyse og konsentrasjon, hvis flere heterozygote bånd må renses igjen etter dialyse og konsentrasjon.
16、Dialyse: legg prøven inn i dialysemembranen, klem begge ender, dialyse ved 4 ℃ over natten;
17、 Konsentrasjon: i henhold til molekylvektstørrelsen til prøven for å velge riktig konsentrasjonsrør 4 ℃ lavhastighetskonsentrasjon, konsentrasjon av proteinkonsentrasjonsmåling, merking, i flytende nitrogen hurtigfrysing, og deretter frosset i -80 ℃ kjøleskap.
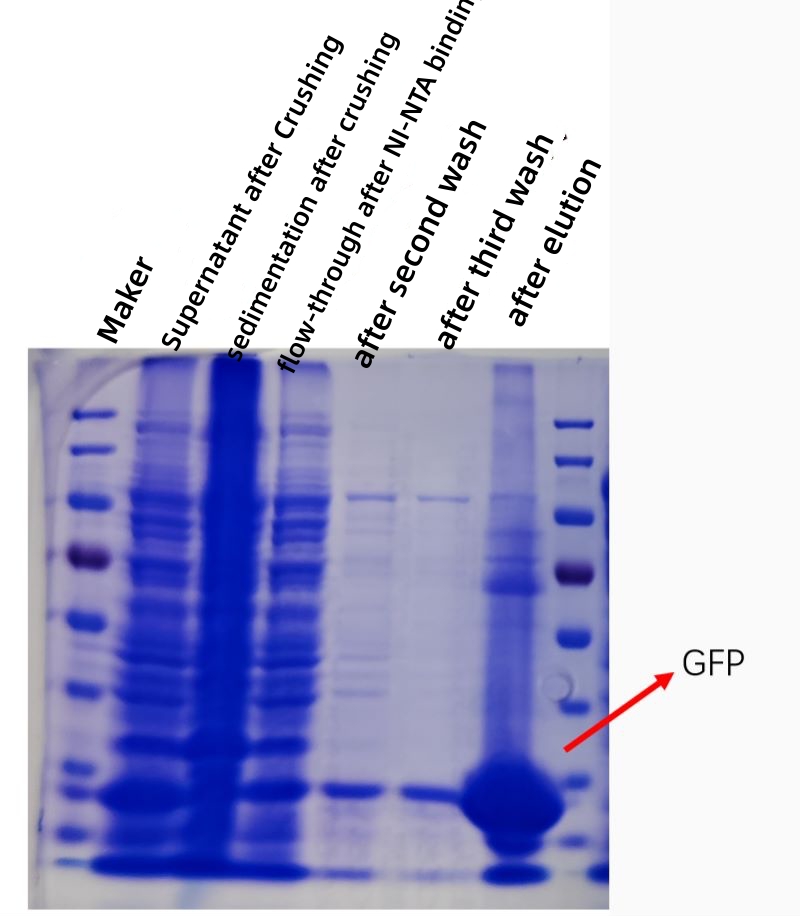
RESULTAT
Figuren nedenfor viser et kofeksjonsplott av GFP-proteinaffinitetsrensing in vitro, og renheten og konsentrasjonen til det rensede proteinet er kraftig økt