Purifikace proteinů je proces zavedení sekvencí nukleových kyselin kódujících proteiny do hostitelských buněk pomocí technik genetického inženýrství, které způsobí jejich expresi ve velkých množstvích, a jejich následné čištění in vitro pomocí vhodných purifikačních metod za účelem získání proteinů vysoké čistoty, aktivity a výtěžku . Proteinové expresní systémy lze rozdělit na prokaryotické systémy a eukaryotické systémy.
Principy a způsoby pro indukci proteinové exprese v prokaryotických systémech a čištění proteinů in vitro pomocí afinitní chromatografie jsou popsány níže.
PRINCIP
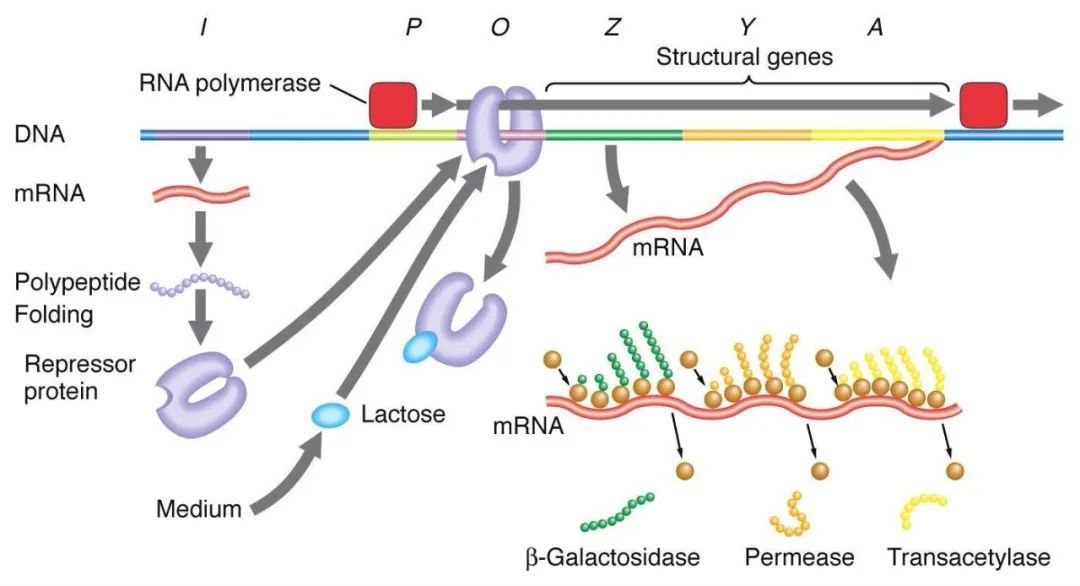
1, Negativní regulace manipulátoru laktózy:
V nepřítomnosti laktózy je lac manipulátor ve stavu odrazení, kdy I sekvence exprimuje Lac odrazující protein, aby se navázal na O sekvenci, čímž zabrání RNA polymeráze ve vazbě na P sekvenci a inhibuje iniciaci transkripce. Když je přítomna laktóza, laktóza vstupuje do buňky a je katalyzována β-galaktosidázou a přeměněna na izolaktózu, která se váže na inhibiční protein, což způsobuje konformační změnu v proteinu a vede k disociaci inhibičního proteinu z O sekvence, čímž se iniciuje transkripce.
Isopropylthiogalaktosid (IPTG) působí stejným způsobem jako isogalaktóza a je extrémně silným induktorem, který není metabolizován bakteriemi a je velmi stabilní, a proto je široce používán v laboratořích.
2, výběr hostitelského kmene:
(1) BL21 je jedním z nejčastěji používaných kmenů pro prokaryotickou expresi, který je vhodný především pro expresi netoxických proteinů pomocí E. coli polymerázy, lze jej tedy aplikovat na expresi prokaryotických systémů pomocí E. coli RNA polymerázy jako je tac nebo trc (např. pGEX, pMAL plazmidy).
(2) BL21(DE3) integruje gen T7 fágové RNA polymerázy v oblasti λ fága DE3 na chromozomu kmene BL21, který může exprimovat jak T7 RNA polymerázu, tak E. coli RNA polymerázu , a může být použit pro expresi plazmidů, jako je řada pET, pGEX, pMAL a tak dále.
3, metody čištění proteinů:
(1) Gelová filtrační chromatografie: podle velikosti molekul od směsi k separaci proteinů, tvaru různých proteinů a molekulární velikosti rozdílů ve směsi přes gelovou filtrační chromatografickou kolonu obsahující částice plniva, kvůli molekulární velikosti různých proteinů jsou různé, difúze do specifické velikosti aperturových částic se schopností měnit se, čím větší budou pozdější molekuly proteinu první, které budou eluovány jako první, tím menší molekuly budou eluovány.
(2) Iontoměničová chromatografie: separace a čištění proteinů je založeno na různých nábojích na povrchu proteinu, povrch proteinu je obvykle nabitý rovnoměrně a za určitých podmínek může být kombinován s katexovými/aniontovými kolonami. Při změně pH nebo použití pufru s postupně se zvyšující iontovou silou k eluci lze navázanou látku vyměnit za ionty v eluentu a eluovat do roztoku. Protože různé látky mají různé náboje a různé vazebné schopnosti s iontoměničovou kolonou, pořadí eluce do roztoku je také odlišné.
(3) Hydrofobní chromatografie: S využitím hydrofobnosti proteinů budou hydrofobní zbytky exponovány na povrchu proteinů po denaturaci nebo v prostředí s vysokým obsahem solí, hydrofobní zbytky různých proteinů mají různou sílu působení s hydrofobními ligandy stacionární fáze a hydrofobnost může být použita jako separace od nejslabší po nejsilnější iónovou složku.
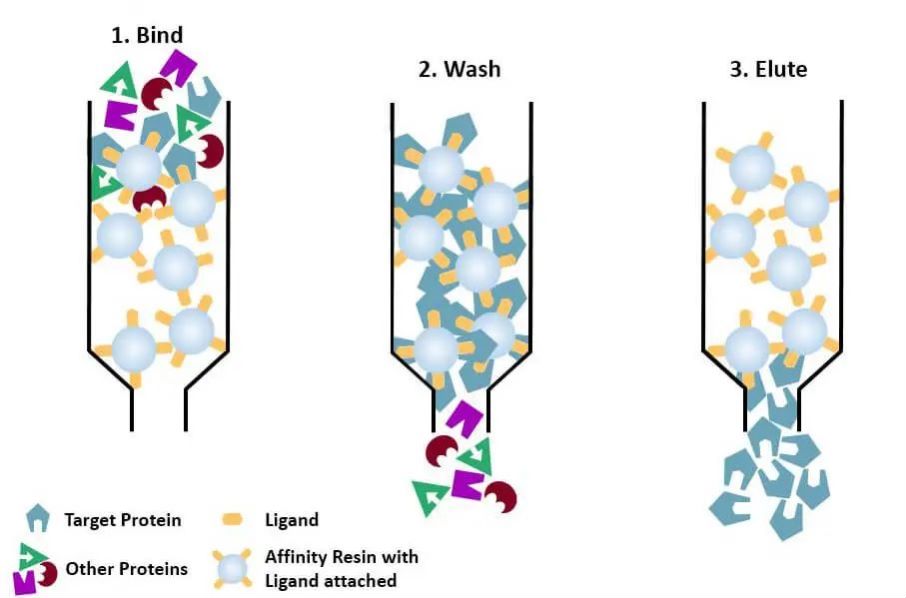
(4) Afinitní chromatografie: specifický ligand proteinu, který má být purifikován (nebo označen na proteinu), je kovalentně připojen k molekule nosiče vhodnými chemickými metodami. Když je proteinová směs přidána do chromatografické kolony naplněné afinitním médiem, protein, který má být purifikován, je specificky navázán na ligand, zatímco ostatní proteiny nejsou navázány a odstraněny promytím a specificky vázaný protein může být eluován roztokem volného odpovídajícího ligandu. Specificky vázané proteiny mohou být eluovány roztokem obsahujícím volný odpovídající ligand.
4、Výběr značek pro čištění proteinů:
Charakteristiky různých štítků jsou následující↓
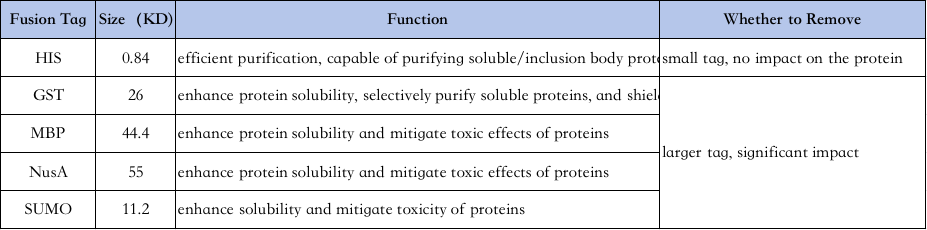
Purifikace His-tag: His-tag je jednou z nejběžněji používaných značek, kde je 6-10 histidinu přidáno na amino nebo karboxylový konec proteinu a cílový protein je purifikován svou schopností pevně se vázat na Ni2+ chelatační kolony za normálních nebo denaturačních podmínek (např. 8M močovina) a poté eluován imidazolem (který soutěží s niklovými jamkami o navázání).
Nástroje a spotřební materiál
Laboratorní činidla:
IPTG; LB médium;
Lyzační pufr: 20 mM Tris (7,9), 500 mM NaCl, 10 mM imidazol;
Promývací pufr: 20 mM Tris (7,9), 500 mM NaCl, 20 mM imidazol;
Eluční pufr: 20 mM Tris (7,9), 200 mM NaCl, 300 mM imidazol ( koncentrace imidazolu může být upravena podle účinnosti eluce );
Dialyzační roztok: 20 mM Tris (7,9), 50 mM NaCl, 10 % glycerol;
Barvicí roztok Kaomas Brilliant Blue; odbarvovací roztok
Nástroje:
Ni-NTA, ultrazvukový přerušovač, chromatografická kolona, koncentrační zkumavky

Postup
1、Konstrukce prokaryotického expresního plazmidu: PCR cílového genu, štěpení vektoru, ligace, transformace, výběr jednoho klonu pro sekvenování;
2、Transformujte úspěšně zkonstruovaný plazmid na BL21(DE3), inkubujte při 37 °C přes noc a vyberte jeden klon pro malé protřepání přes noc;
3、Malá indukce pro zjištění nejlepších podmínek indukce: bude třepán přes noc bakteriální roztok 1:1000 inokulace do 3mL LB, 37 ℃ třepání 4-5h do bakteriálního roztoku OD600 = 0,6-0,8, přidat různé koncentrace IPTG (0,1-1mM se prodlužuje doba, např. při různých teplotách), např. 37 ℃ indukované 4-5h, 30 ℃ indukované 6h-8h, 16 ℃ indukované 16-20h, 16°C indukované 16-20h, teplota se sníží, doba se prodlouží, např. 37 ℃ indukované 4-5h, 30 ℃ 6h °C indukované 16-20h, teplota se sníží. C po dobu 16-20h, odeberte bakteriální mízu před a po indukci za různých indukčních podmínek, proveďte gel, obarvte a sledujte výsledky indukce; (Obecně řečeno, čím nižší je koncentrace IPTG a čím nižší je indukční teplota, tím pomalejší je exprese cílového proteinu, tím více přispívá ke správnému skládání proteinů, čímž se zvyšuje jejich rozpustnost a snižuje se tvorba inkluzních tělísek)
4、Po nalezení nejvhodnějších podmínek indukce naočkujte bakteriální roztok v poměru 1:1000 do 2 l LB, protřepejte při 37 °C, dokud OD600 bakteriálního roztoku = 0,6-0,8, aspirujte 20 μl bakteriálního roztoku a nechte jej proběhnout gelem (1), poté navodte vhodné podmínky indukce a indukujte získané proteinové exprese za podmínek exprese 20 μl bakteriálního roztoku a nechte na něm působit gel (2);
5、Odstraňte indukovanou bakteriální kapalinu, centrifugujte při 4000 g po dobu 15 minut při 4 °C;
6、Zlikvidujte supernatant, zvažte, přidejte 10 ml lyzačního pufru (1:100 plus inhibitor proteázy) na každý gram bakterií, resuspendujte, umístěte na led po dobu asi 30 minut;
7、Drcení pod tlakem: odložte alkohol do vysokotlakého homogenizátoru, dvakrát opláchněte vodou, jednou použijte lyzační pufr k vyvážení, přidejte bakteriální kapalinu, natlakujte (tlak by neměl překročit 800 kpa), bakteriální kapalinu třikrát až pětkrát, aby byla průhledná a nelepivá;
8、Shromážděte rozdrcenou bakteriální kapalinu, 12000 g, 4 °C odstřeďování po dobu 20 minut, separace supernatantu a sraženiny, každý zadržený 20 μl vzorku (3) (4), který má být spuštěn gel;
9,2 ml Ni-NTA bylo přidáno do purifikační kolony, aby byla filtrována ethanolem, opláchnuta vodou a přidán lyzační pufr pro ekvilibraci kolony;
10、S resuspenzí lyzačního pufru Ni-NTA přidanou k supernatantu, dobře promíchejte, inkubace na třepačce při 4 °C po dobu 2 hodin;
11. Supernatant prošel kolonou při 4 °C a shromáždilo se 20 ul filtrátu (5);
12、Promyjte kolonu 5mL promývacího pufru 3x, odeberte vzorek filtrátu 20μL (6);
13、Přidejte 1 ml elučního pufru, inkubujte 5 minut, odeberte eluát, opakujte 5krát, odeberte 5 ml eluátu do stejné zkumavky, ponechte 20 μl vzorku (7);
14、Jednotlivé proteinové vzorky získané v průběhu experimentu byly analyzovány, obarveny Caumas Brilliant Blue po dobu 1 hodiny a odbarveny, dokud nebyla modrá barva pozadí světlejší a nebyly viditelné jasné proteinové pásy;
15、Analyzujte proteinové pásy každého vzorku podle výsledků následné operace: pokud eluované proteinové vzorky v proteinu bez heterozygotních pásů nebo heterozygotních pásů nebo méně a mělké, lze provést dialýzu a koncentraci, pokud je třeba po dialýze a koncentraci znovu purifikovat více heterozygotních pásů.
16、Dialýza: přidejte vzorek do dialyzační membrány, upněte oba konce, dialýzu při 4 °C přes noc;
17、Koncentrace: podle velikosti molekulové hmotnosti vzorku vyberte vhodnou koncentrační zkumavku 4 ℃ nízkorychlostní koncentraci, měření koncentrace proteinu, značení, rychlé zmrazení v tekutém dusíku a poté zmrazení v chladničce -80 ℃.
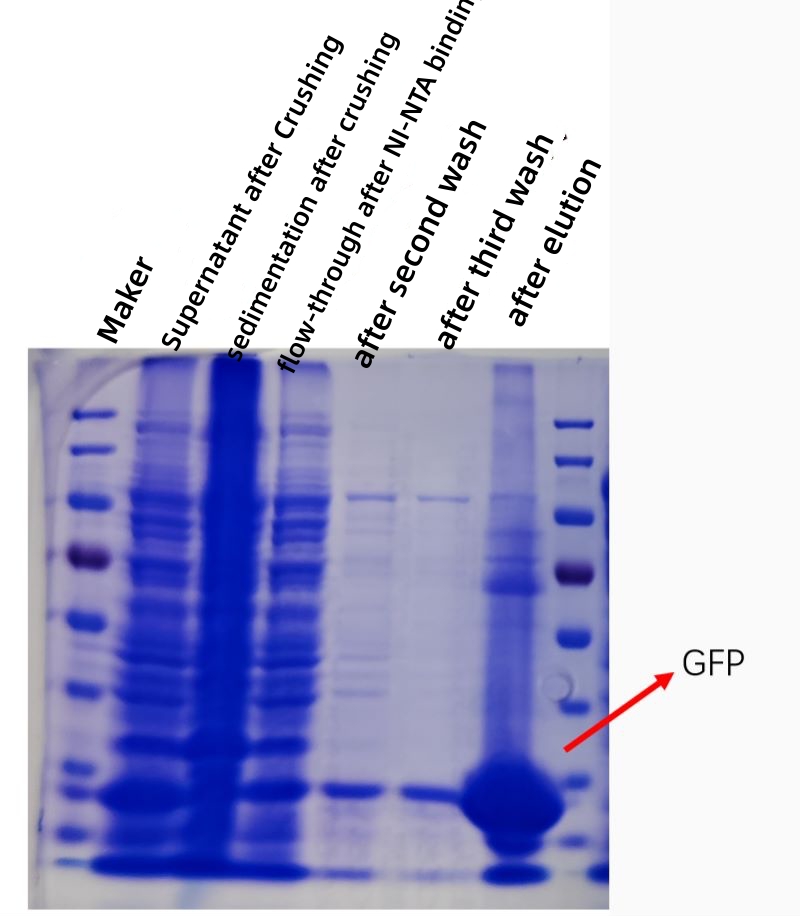
VÝSLEDEK
Obrázek níže ukazuje kofekční graf afinitní purifikace GFP proteinu in vitro a čistota a koncentrace purifikovaného proteinu je značně zvýšena